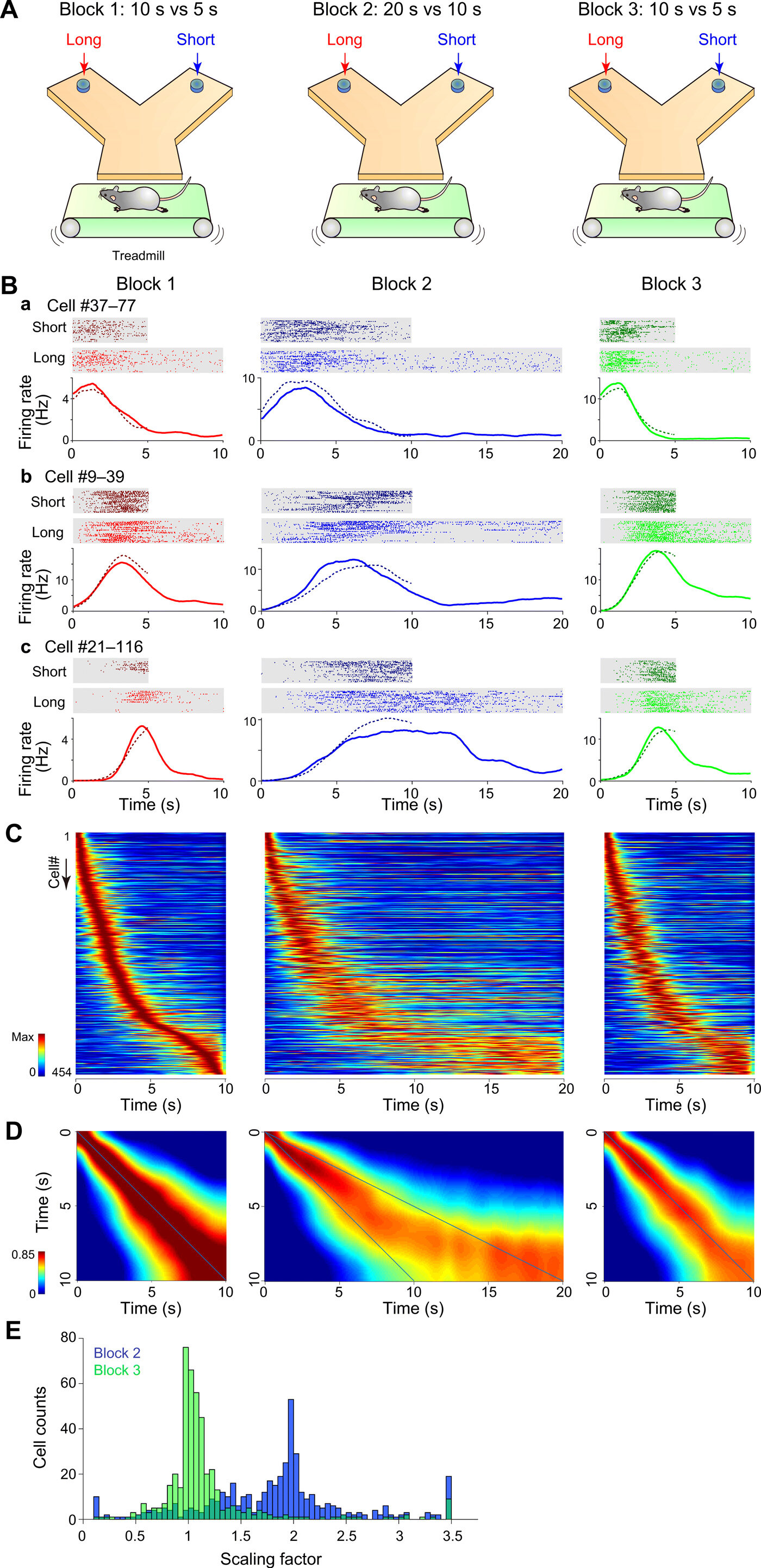
Клетки времени гиппокампа могут кодировать определенные моменты организованного опыта во времени, чтобы поддерживать функции гиппокампа эпизодической памяти. Но мало что известно о перестройке временных ячеек при своевременной смене эпизодов. Акихиро Симбо и исследовательская группа из центра науки о мозге Рикена и факультета психологии Университета Кейо в Японии исследовали активность нейронов CA1, которые имеют решающее значение для автобиографической памяти, ментальных путешествий во времени и аутоноэтического сознания у людей. Во время выполнения задач временного деления они проектировали наборы временных интервалов, которые должны быть расширены, дискриминированы или сокращены по блокам испытаний. Команда масштабировала прецессию тета-фазы и тета-последовательности временных ячеек, чтобы сохранить тонкие временные отношения между парами в тета-циклах. Используя модели крыс, они показали, как тета-последовательности отражают решения о ставках на основе их времени. Полученные результаты показали, как масштабируемые функции временных ячеек могут поддерживать гибкие временные представления формирования памяти.
Когнитивные карты
Нейробиологи предлагают животным иметь когнитивную карту, которая служит внутренним представлением отношений между сущностями в мире и поддерживает гибкое поведение. Гиппокамп может предоставлять когнитивные карты для пространственной навигации, поскольку сборки клеток гиппокампа могут организовывать картографические представления окружающей среды. Клетки Гиппокампа используют как темпоральное, так и темпоральное (временное) кодирование для представления пространства. Кодирование скорости принимает форму рецептивных полей локаций. Спайковые тайминги ячеек места могут быть сильно модулированы параллельными тета-колебаниями от 4 до 10 Гц, проявляя при этом спайковые фазовые сдвиги в зависимости от расстояния относительно центра рецептивного поля. При этих двух формах темпорального и темпорального кодирования пространственные представления гиппокампа включали как позиционную, так и реляционную информацию. Относительные позиционные отношения могут обеспечить более фундаментальную информацию для пространственного распознавания в гиппокампе, чем абсолютные положения. Гиппокамп также обеспечивает когнитивную карту для внепространственных сущностей, включая информацию, основанную на времени. Однако не так много исследований посвящено изучению систематической реорганизации представлений о времени в гиппокампе. В этой работе Shimbo et al. изучал масштабируемость времени в гиппокампе. Гиппокамп представляет информацию о прошедшем времени последовательной активностью временных клеток. В предыдущих докладах подробно описывалось, как несколько областей мозга, включая префронтальную кору и средний мозг, участвуют в восприятии и представлении прошедшего времени.
Эксперименты
Во время работы исследователи выполняли задания на деление пополам для крыс, чтобы различать длинные и короткие интервалы, чтобы получить вознаграждение. Они отметили, что большинство временных ячеек в области CA1 представляют временную информацию в масштабе пробных блоков. Shimbo et al. исследованы временные структуры последовательности активности временных клеток, связанные с тета-колебаниями в гиппокампе, с целью выявления механизмов, лежащих в основе масштабируемого представления времени. Анализируя полученные данные, они отметили активность временных клеток, отражающих решения крыс на основе их восприятия времени. Полученные результаты показали общий механизм схемы, лежащий в основе картографических представлений пространственной и внепространственной информации в гиппокампе из-за сходства между клетками места и клетками времени. Ученые использовали кремниевые зонды высокой плотности для записи информации от нейронов CA1 гиппокампа крыс во время выполнения задания. Они классифицировали единицы как предполагаемые пирамидальные клетки или интернейроны на основе их формы волны и скорости стрельбы.
Масштабируемое отображение времени
Shimbo и соавт. обученных крыс на временное деление пополам задач, и установить временные интервалы, чтобы различать блоки испытаний. Во время экспериментов они заставляли крыс бегать на беговой дорожке в течение длительного или короткого времени. После вынужденного бега они выбирали левую или правую руку в Y-образном лабиринте. Исследователи предоставляли капли воды в качестве награды при выполнении каждого задания на правильной руке. Они впервые оценили зависящую от времени активность пирамидных клеток и классифицировали 22,2% пирамидных клеток как клетки времени. Они также определили, масштабируется ли временное представление пирамидальных клеток CA1 в направлении сжатия—чтобы указать, что временное представление пирамидальных клеток CA1 масштабируется как в направлении расширения, так и в направлении сжатия.
Зависимость задачи от масштабируемого представления времени
Чтобы проверить, как масштабируемые характеристики времени также зависят от требований задач, Shimbo et al. обучили другую группу крыс задачам легкой дискриминации, в которых животные не должны были различать временные интервалы. Во время выполнения заданий они заставляли крыс бегать по беговой дорожке с короткими или длинными интервалами, после чего требовали, чтобы животные выбирали левое или правое крыло Y-лабиринта, ориентируясь на свет. Затем они исследовали активность нейронов CA1 у крыс, выполняющих задачи световой дискриминации. Исследователи классифицировали 7,5% пирамидных клеток как временные клетки, которые были значительно ниже, чем те, которые были получены в задачах временного деления пополам. Исходя из результата, формирование временных ячеек зависело от требований задачи, а также от масштабируемого представления информации о прошедшем времени.
Масштабируемость тета-фазовой прецессии и тета-последовательности временных ячеек
Сборки клеток гиппокампа генерировали последовательные структуры во время космической навигации в одиночных тета-циклах, известных как тета-последовательности. Эти циклы компактно представляли собой траекторные последовательности прошлых, текущих и будущих позиций. Shimbo et al. проверено наличие прецессии тета-фазы в зависимости от прошедшего времени во время выполнения задачи временного деления пополам. В локальном потенциале поля преобладали устойчивые колебания тета-частоты (от 4 до 10 Гц) в течение интервального периода. Команда изучила фазово-временные отношения отдельных нейронов и обнаружила, что 77,3% временных клеток представляют собой фазовую прецессию. Shimbo et al. исследовал временную структуру спайковых последовательностей сборок временных ячеек в одиночных тета-циклах, чтобы понять, формировали ли сборки временных ячеек тета-последовательности во время выполнения задачи.
Результаты представлены временными отношениями пар клеток в одиночных тета-циклах. Затем команда оценила, были ли тета-последовательности временных клеток просто следствием фазовой прецессии отдельных нейронов или они имели более высокие корреляционные структуры, чем те, которые предсказывались из фазовой прецессии. Результаты показали, что тета-последовательности временных клеток были не просто следствием фазовой прецессии отдельных нейронов. Вместе эти результаты показали, как тета-последовательность временных ячеек масштабируется по всем испытательным блокам и как тонкие временные отношения между клеточными парами сохраняются в пределах одного тета-цикла. Ученые также изучили, как сборка временных клеток отражала распознавание прошедшего времени у крыс. Работа показала, как эти результаты указывают на активность временных клеточных агрегатов и фактически отражают решения животных во время тестовых испытаний.
Прогноз
Таким образом, Акихиро Симбо и его коллеги исследовали масштабируемость временных клеток в гиппокампе относительно скорости и временного кодирования. Результаты показали масштабируемость временного представления в гиппокампе и пирамидных клетках гиппокампа относительно двух факторов-скорости и времени. В этой работе предлагается лежащий в основе общий контурный механизм с картографическими представлениями пространственной и внепространственной информации в гиппокампе. Такие масштабируемые, картографические архитектуры пространственно-временной информации позволят сформировать гибкую память для адаптации к сложным средам. | |
| Просмотров: 365 | |